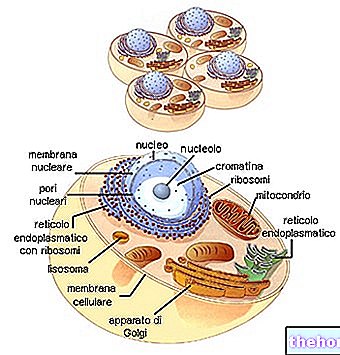
Цикличната приемственост на живите същества намира връзките между следващите поколения във феномените на възпроизводството.
Възпроизвеждането се извършва на различни нива от еволюционната скала, в различните разклонения на растителното и животинското царство, в различните живи видове, с такова разнообразие от механизми, които единствено оправдават цял трактат.
Първата класификация на явленията на размножаване трябва да разграничава едноклетъчните от многоклетъчните организми, тъй като само в първите клетъчното делене съвпада с размножаването.
При многоклетъчното размножаване може да бъде агамно или сексуално (или гамично).
Агамичното размножаване, сравнително по -рядко, се основава на механизма на митозата, така че променливостта на вида е по -скоро поверена на многократната поява на мутации.
Съществуват и различни механизми, като стробилизация, спороношение и т.н., докато в растенията откриваме регенеративни форми, добре познати в селското стопанство (резници, наслояване и т.н.).
Най -разпространеният репродуктивен механизъм при висшите форми обаче е половият, съответстващ на появата на мейоза, образуването на гамети и сливането им в зиготата (оплождане).
При примитивните видове гаметите не са морфологично диференцирани: в този случай говорим за изогаметия. Винаги обаче има две серии гамети, идентифицирани със символите (+) и (-) и оплождането може да се осъществи само от „срещата между гамети с противоположен знак: следователно има биологична разлика, която все още не е изразена морфологично.
С напредването на еволюционната скала се появява морфологична и функционална диференциация, при която тип женски гамети обикновено са снабдени с изобилие от резервен материал (девтоплазма или теле, което ще осигури развитието на ембриона, докато не стане метаболитно независим) и вид мъжки гамети, надарени с подвижност, за да достигнат женски. Гаметите винаги са хаплоидни и са резултат от мейоза. Сливането им поражда диплоидната зигота.
Между мейозата и зиготата може да изтече поредица от хаплоидни клетъчни поколения, точно както между зиготата и мейозата може да изтече поредица от диплоидни клетъчни поколения, като различните разновидности на генеративните цикли се изразяват чрез редуването на поколенията.
Диплонтен вид (с диплоиден организъм) се характеризира с гаметична мейоза: мейозата директно произвежда гаметите, които чрез сливане незабавно възстановяват диплоидното състояние. Това е преобладаващият случай на метазоанците, включително човека.
Гаметогенеза
Като се има предвид възпроизводството с гаметична мейоза, както се среща при човека, нека се опитаме да изясним как мейозата се вписва в гаметогенезата (образуването на гамети).
В ембрионалното развитие, на мъжката и женската гаметогенеза (наречена сперматогенеза и оогенеза), има ранна диференциация между клетките, предназначени да образуват тялото (соматична линия), и тези, предназначени да произвеждат гамети (зародишна линия). Първоначалните клетки на зародишната линия се наричат протогони. С диференциацията на гонадите в мъжкия или женския смисъл има диференциация на зародишните клетки съответно в сперматогония и овогония.
Разглеждайки сперматогенезата, виждаме, че в сперматогонията има поредица от клетъчни поколения, която продължава през целия живот. Само част от така непрекъснато произвежданата сперматогония се различава от нормалния митотичен цикъл и вместо него започва мейотичният.
Зародишната клетка, в която ще започне мейозата (редупликация и след това първо деление), се нарича сперматоцит от първи ред; разделянето му поражда два сперматоцита от втори ред, които с второто разделение пораждат общо четири сперматиди.
Можем да завършим редукцията на хромозомния комплект от 4n от сперматоцити от първи ред (след редупликация има четири хроматила за всяка двойка хомолози) до 2n от сперматоцити от втори ред и до n от сперматидите, както вече се вижда при изучаването мейоза, която Така заключаваме. Следователно сперматидите вече са хаплоидни, но все още не са зрели гамети. От структурата на клетка от хаплоиден тип, функционалното съзряване (наречено спермиохистогенеза) трансформира сперматидите в сперматозоиди, тоест зрели мъжки гамети.
В женската гаметогенеза (или оогенеза) има няколко разлики. На първо място, броят на подготвените гамети е много по -малък. Смята се, че около 5 X 105 овогони се приготвят в половите жлези на женска от човешкия вид; от тях само около 400 се интересуват от узряване на фоликулите и последващо изтласкване, в цикъл, който обикновено засяга само един фоликул на месец за период на плодородие от около 35 години.
Различният брой гамети, приготвени при двата пола, съответства на вече споменатата разлика във функцията и поведението: сперматозоидите са малки, подвижни и многобройни във връзка с необходимостта от търсене на яйцеклетката и ниската вероятност да я намерят; яйцата са големи, инертни и малко, във връзка с функцията да гарантират на ембриона резервния материал и защитата, която им се осигурява чрез вътрешно оплождане (естествено, особено при външно оплождане, яйцеклетките също трябва да са по -многобройни).
Необходимостта от осигуряване на гамети с резервен материал съответства на наличието в "оогенезата" на фаза на спиране на мейозата, по време на която хромозомите са частично деспирализирани. След това наблюдаваме така наречените "перо хромозоми", в които серия на екстроверсиите идентифицира чертите, при които гените, отговорни за синтеза на девтоплазма, са депресирани.
По -малкият брой, необходим за женските гамети, също съответства на факта, че от четирите хаплоидни клетки, произведени от мейозата, само една получава целия резервен материал и се превръща в гамета, докато другите три (полоцитите или полярните тела), които съдържат само хромозомни материали, не могат да дадат началото на зиготи и ембриони и са предназначени да регресират.
Оплождане
Оплождането, тоест срещата между мъжката и женската гамета, може да се извърши по много различни начини.В животинското царство наблюдаваме прехода от външно оплождане (гамети, изложени на всякакъв екологичен риск и следователно непременно многобройни при двата пола) към вътрешно оплождане, с което родителските грижи са допълнително свързани с метаболитната връзка на бозайниците между майката и плода .
Оплождането, след като срещата между гамети от противоположния пол е настъпила, трябва да се осъществи, гарантирайки две условия: специфичност и уникалност. Тоест трябва да се гарантира, че сперматозоидът е от същия вид като яйцеклетката и че след като влезе първият, няма да влязат други.
Специфичността се осигурява от биохимичните характеристики на акрозомата и повърхността на яйцеклетката. Всъщност се говори за реакции между «фертилизини» и «антифертилизини», със специфичност, сравнима с тази на срещата между ензими и субстрат.
Уникалността на оплождането е гарантирана от модификация на повърхностната структура на яйцеклетката („кортикална реакция“), която започва точно в момента на първата специфична реакция фертилизин / антифертилизин; след тази реакция мембраната на яйцеклетката се променя, така че всички други сперматозоиди, които достигат до нея, вече не са в състояние да инициират специфичната реакция на оплождане.
След оплождането опашката на сперматозоида остава външна за яйцеклетката, докато хромозомният материал прониква в нея. Това, наречено "мъжки пронуклеус", се присъединява към "женския пронуклеус" на яйцеклетката, като по този начин образува диплоидното ядро на зиготата.